毛状体是覆盖大多数陆生植物地上样本的特殊突起。不同种类的毛状体在物种中存在很大的差异,因此大多数植物物种中毛状体形成的分子调控机制和生物学功能仍未被探索。2024年9月4日,美国内布拉斯加大学林肯分校的Joe Louis教授团队在New Phytologist期刊上发表了题为“Impaired Brown midrib12 function orchestrates sorghum resistance to aphids via an auxin conjugate indole‐3‐acetic acid–aspartic acid”的研究论文。研究者通过转录组与植物激素检测技术对高粱褐色叶中脉突变(bmr12)材料,过表达材料及野生型材料进行分析,揭示了高粱中褐色叶中脉突变通过调控植物激素吲哚-3-乙酸-天冬氨酸(IAA-Asp)积累,增强了对高粱蚜的抗性。


1.高粱蚜取食调节高粱基因表达及木质素积累
高粱Bmr6和Bmr12基因分别编码CAD酶和COMT酶,CAD酶催化羟基肉桂酰醛转化为单脂醇,COMT酶负责催化单脂醇生物合成途径的倒数第二步。作者检测了高粱蚜(SCA)取食RTx430植株早期基因Bmr6和Bmr12的表达水平。SCA取食24小时后增强了Bmr6和Bmr12基因的表达水平,48小时显著降低(图1)。Western blots检测了Bmr12表达变化相关的Bmr12 (COMT)抗体(图1c)。Mäule染色的组织化学分析显示,SCA取食10天后,RTx430高粱叶片中S-木质素的沉积减少(图1)。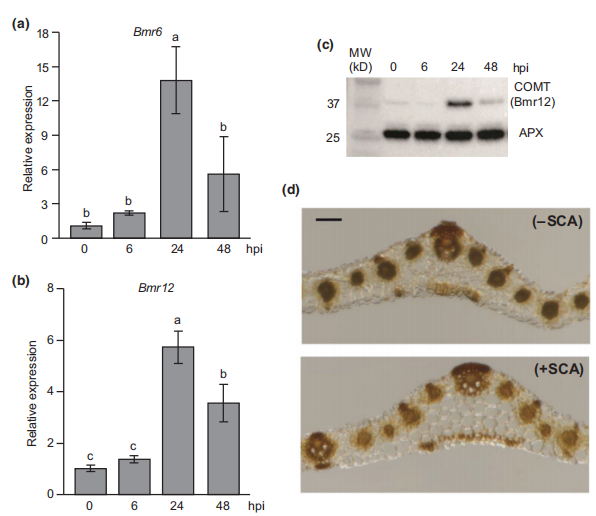
图1 高粱蚜取食后高粱植株基因变化情况
2.Bmr12突变增强高粱对SCA的抵抗力
作者对bmr6、bmr12以及bmr6和bmr12基因的过表达系及突变系进行了无选择实验,以测试单脂醇在提供SCA防御中的作用。与野生型植株相比,SCA取食5和10天后,bmr12突变植株的SCA数量显著降低,bmr12 - oe1植株的SCA数量显著增加(图3b,d)。这些结果表明,Bmr12功能的丧失增强了高粱对SCA的抗性。此外,bmr12敲除系和bmr12 - oe1植物的Mäule染色分别显示s -木质素沉积减少和增强。作者评估了SCA在野生型,bmr12敲除系和bmr12 - oe1植物达到首个筛管元素阶段(SEP)所需的总时间,与野生型和Bmr12-OE1植物相比,SCA在bmr12植物上达到首个SEP所需的时间显著更长。
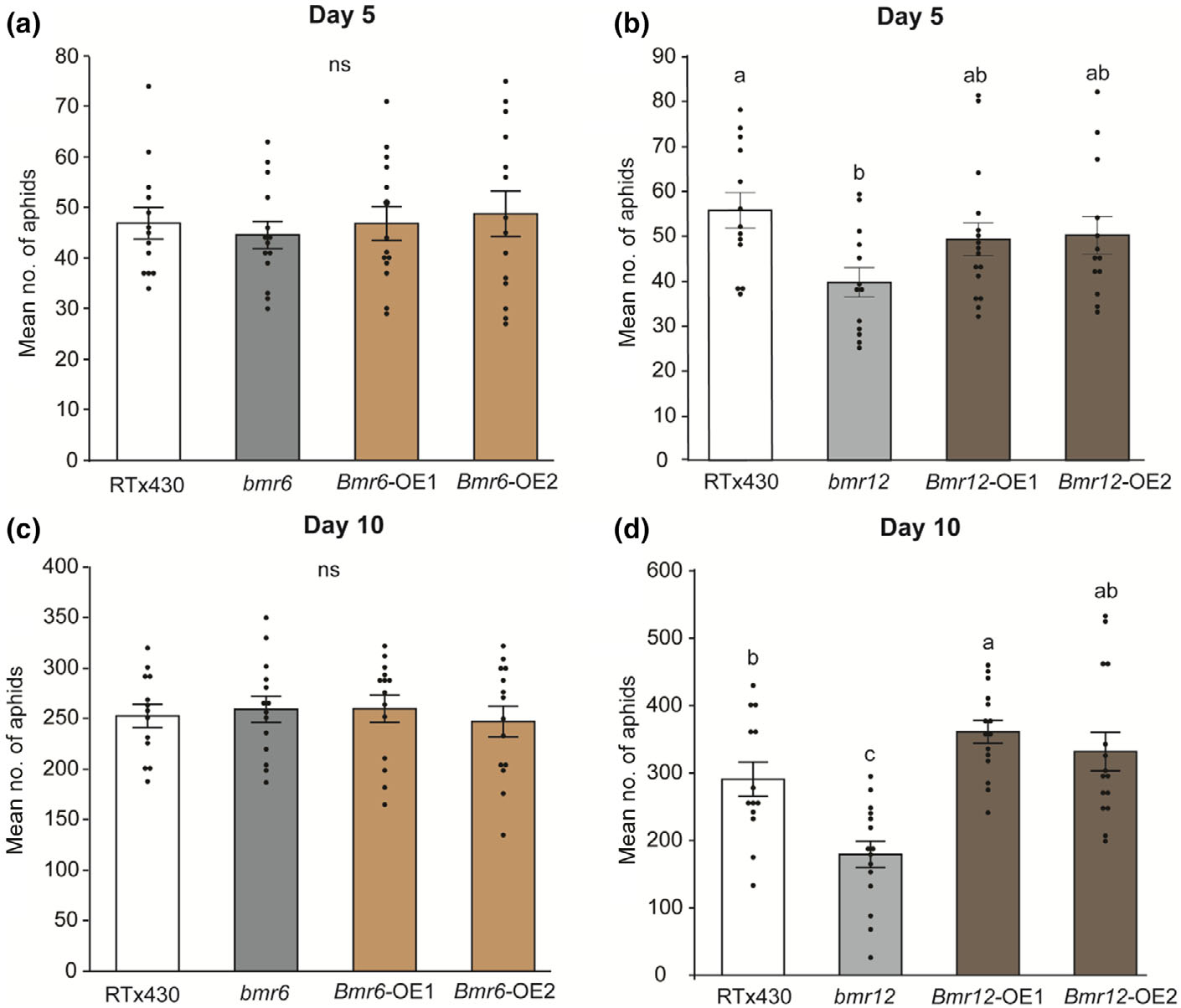
图2 关键基因过表达及敲除实验对高粱蚜取食反应
3.SCA取食改变了高粱转录组
对高粱三个材料(野生型、bmr12敲除系和Bmr12-OE1)进转录组分析,与5天接种后相比,10天接种后的数差异基因数量在所有三个材料中都更高。对5和10 dpi时的差异基因进行了GO富集分析。在10 dpi时,bmr12敲除上调的216个差异基因富含与初级代谢(脂质分解过程、碳水化合物和蔗糖代谢过程)和防御(对生长素、激素和内源刺激的响应)相关的GO术条目。Bmr12-OE1在10 dpi时上调的213个独特基因富含与初级代谢(光合作用和羧酸生物合成过程)、氨基酸生物合成过程以及氧化还原过程相关的GO条目。同样,野生型植物在10 dpi时上调的162个独特基因富含与蛋白质磷酸化、蛋白质代谢过程、磷代谢过程和离子转运相关的GO条目。GO富集分析揭示了只有高粱bmr12敲除系在响应SCA取食时显示出与防御相关的GO条目。
4.JA和SA途径不参与bmr12介导的SCA抗性
在高粱中,通过转录组数据分析发现,与木质素生物合成相关的基因表达水平在SCA取食后发生了变化。特别是在bmr12基因突变体中,与茉莉酸(JA)和水杨酸(SA)途径相关的基因表达模式发生了显著变化,这可能与植物对SCA的抗性有关。在bmr12敲除系中,与SA途径相关的基因表达上调,这表明bmr12可能通过调节SA途径来增强对SCA的抗性。而在Bmr12-OE1品系中,JA-Ile(茉莉酸异亮氨酸)的水平显著升高,但在SCA取食后被抑制,这可能表明JA途径在调节对SCA的抗性中也起着作用(图3)。
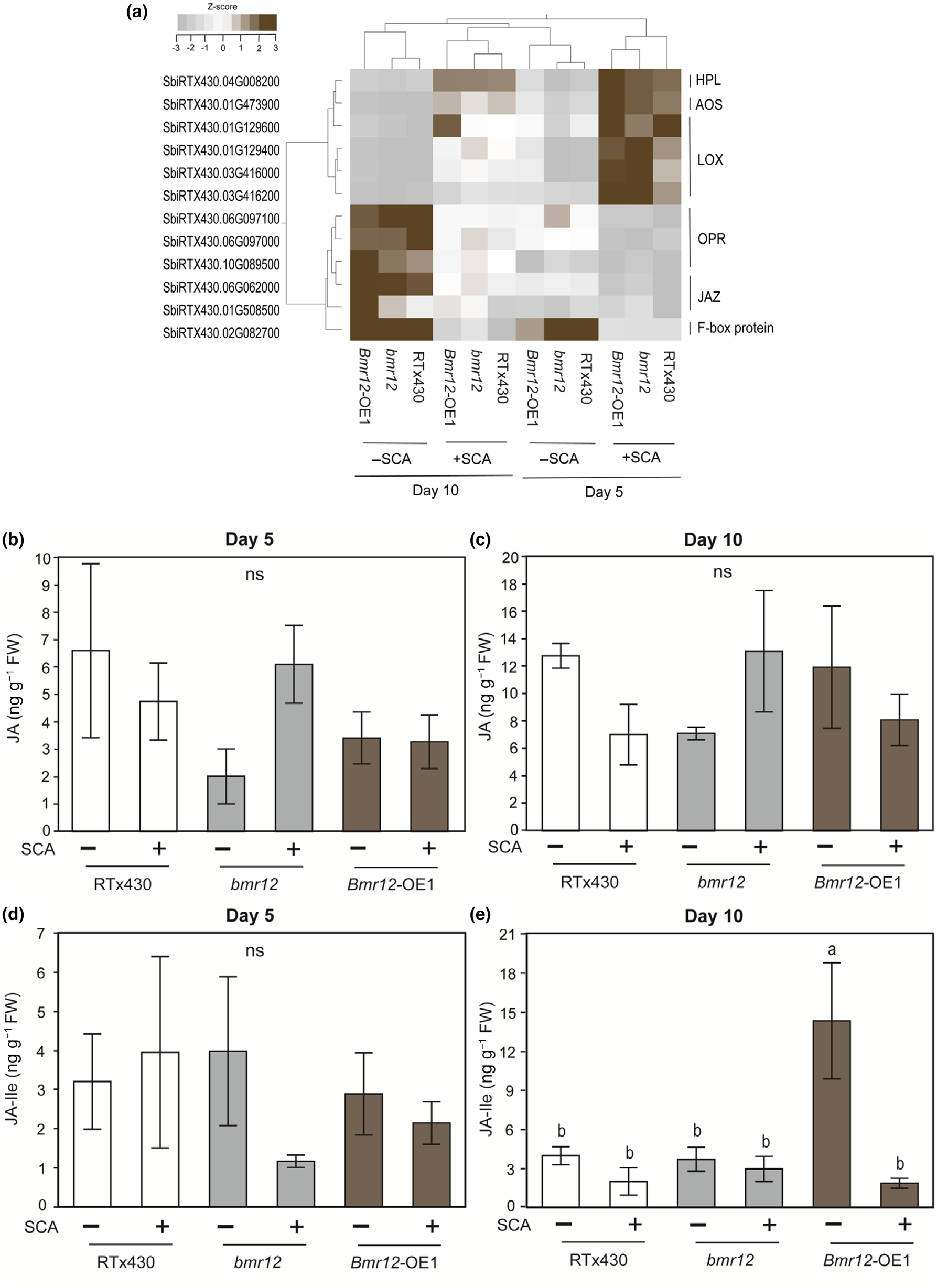
图3 SCA取食对高粱茉莉酸(JA)途径基因及代谢产物的影响
此外,研究发现,SCA取食增加了所有三个高粱材料中SA的水平,但在bmr12敲除和Bmr12-OE1系中,SA水平在SCA取食后并没有显著变化,这表明bmr12介导的对SCA的抗性可能独立于SA途径。而在RTx430品系中,SA水平在10 dpi时显著升高,这可能与该品系对SCA的抗性有关(图4)。
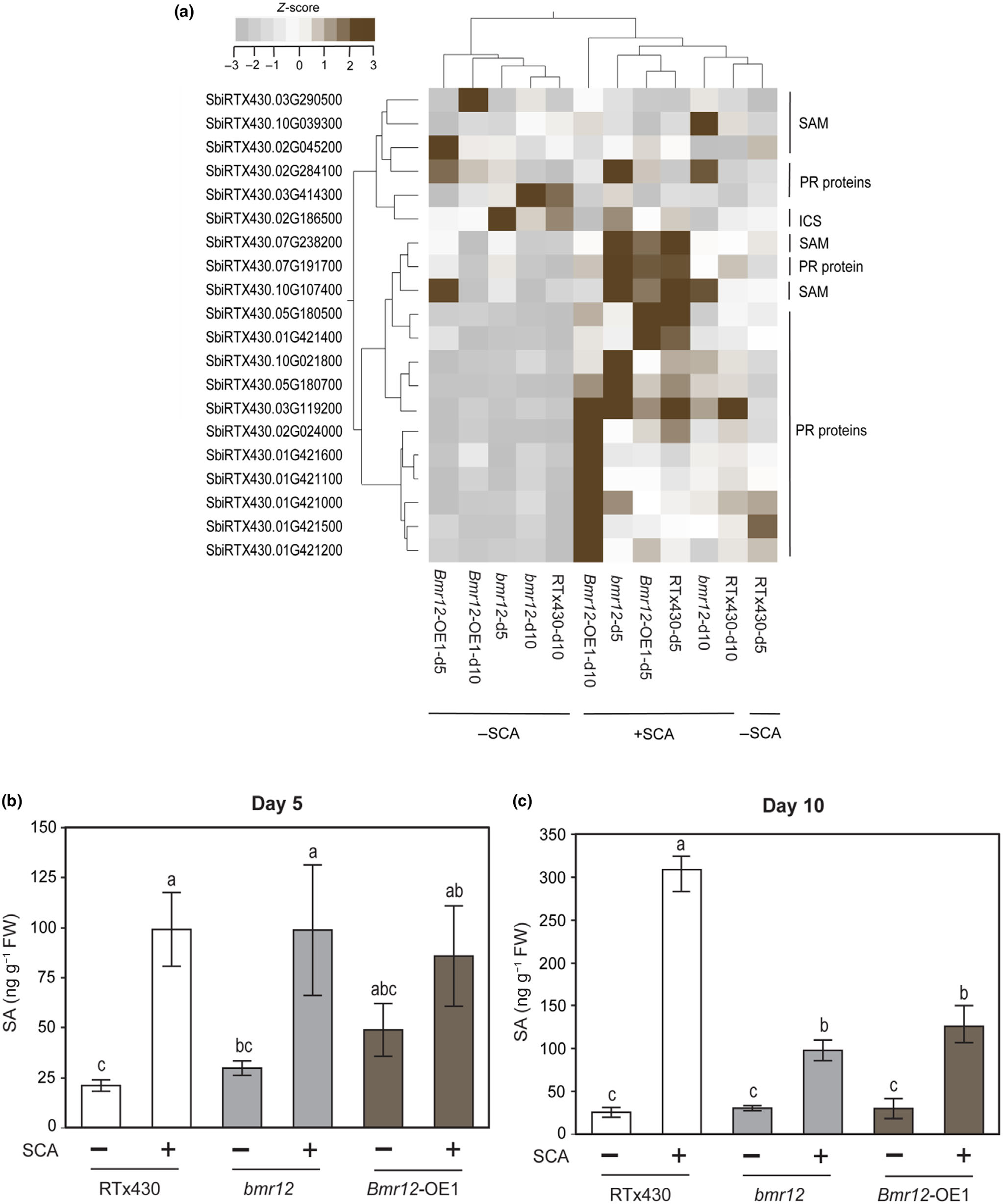
图4 SCA取食对高粱水杨酸(SA)途径基因及代谢物的影响
5.SCA摄食触发bmr12植物生长素代谢和IAA-Asp水平相关基因的表达
生长素应答基因、SAUR-like、GH3家族蛋白等与生长素信号通路相关的几个基因在三个材料中5或10 dpi时均显著上调。两个编码生长素代谢和应答因子的基因在5和10 dpi时显著下调。三个 SAUR-like生长素响应家族蛋白在10 dpi时在bmr12敲除系中显著上调,而在其他材料中未发现变化。总体而言,与生长素代谢相关的基因在bmr12敲除植物中存在差异表达(图5a)。通过植物激素量化,在SCA侵染前后,bmr12敲除系植株在5 dpi和10 dpi的总IAA水平均未观察到显著变化(图5b),但在10 dpi时,RTx430和bmr12 - oe1植株的IAA水平显著下降(图5c)。此外,我们发现5dpi时bmr12敲除系植株的IAA-Asp水平显著升高(图5d)。外源IAA-Asp可恢复Bmr12过表达植株的抗性。
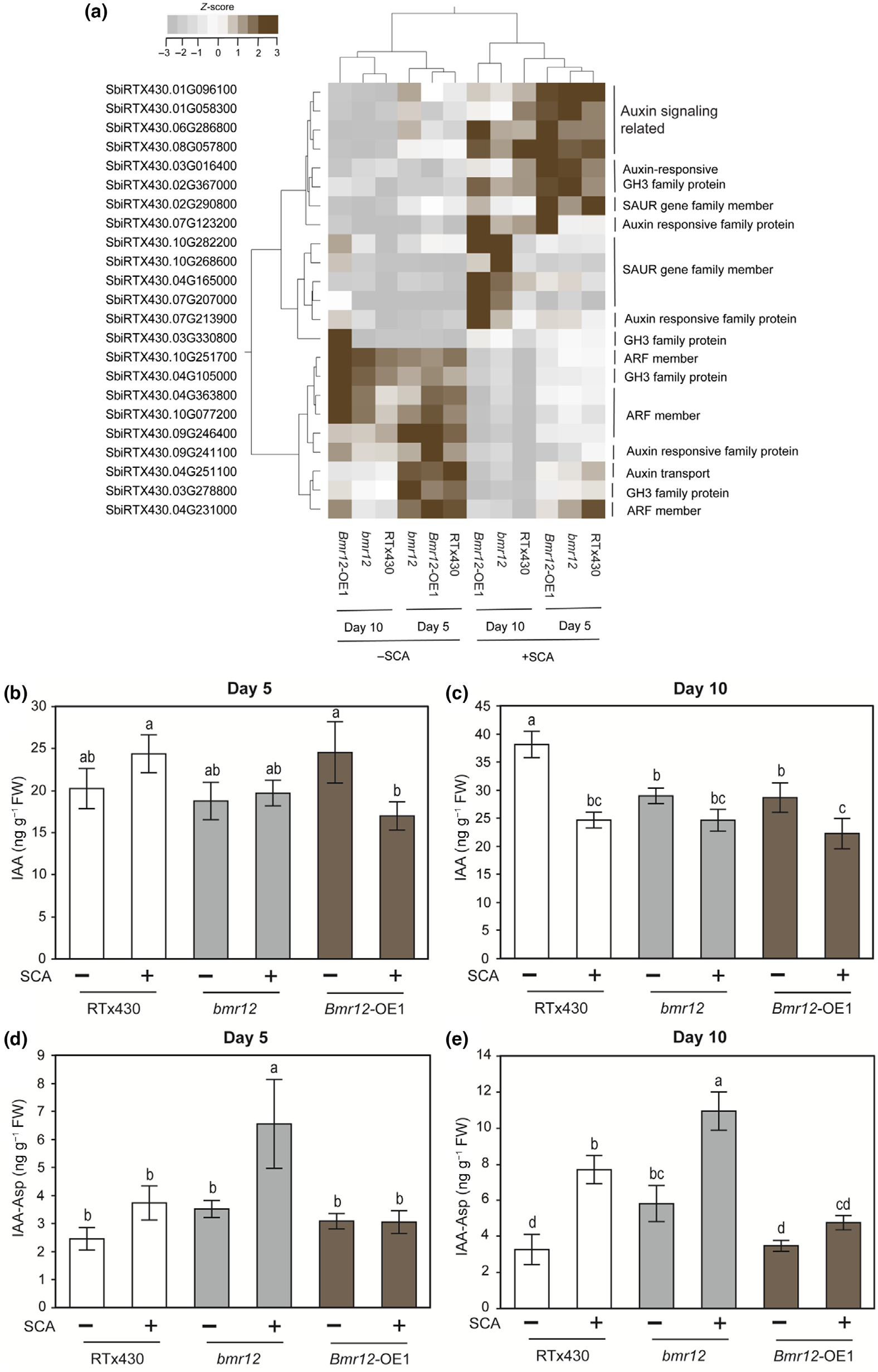
图5 高粱蚜侵染对高粱植物生长素途径基因及代谢产物的影响
为了证实IAA-Asp在高粱抗蚜中的作用,作者在3个高粱材料上外源施用了50 μ m的IAA-Asp。在5和10 dpi时,与对照植株相比,野生型和Bmr12-OE1系上的蚜虫数量减少(图6a,b)。对照和经IAA-Asp预处理的bmr12敲除系植株在5和10 dpi时的SCA数量没有差异(图6a,b),表明外源施用IAA-Asp不会导致对蚜虫抗性的增强。值得注意的是,研究结果表明,与对照植株相比,外源IAA-Asp在5和10 dpi时恢复了Bmr12-OE1植株的抗性(图6a,b)。因此,得出结论,IAA-Asp是bmr12赋予蚜虫抗性的关键成分。
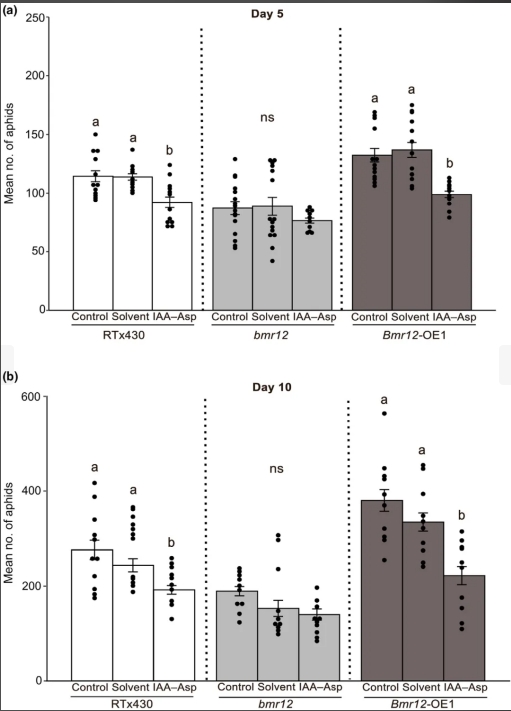
图6 IAA-Asp处理后,3个高粱材料高粱蚜成虫和若虫数量
6.高浓度的IAA-Asp对SCA繁殖有不利影响
最近,有研究认为IAA-Asp可能与提高番茄对紫丁香疫病毒的防御能力有关。此外,由于与Bmr12-OE1对照植株相比,外源IAA-Asp使Bmr12-OE1植株恢复了SCA抗性,因此作者研究了IAA-Asp是否对SCA生长和繁殖力有直接的负面影响。为了评估这一点,将SCA饲养在含有不同浓度IAA-Asp的人工饲料中3天。蚜虫饲养试验表明,与对照组相比,人工饲料中添加高浓度(25、50和100 lM)的IAA-Asp导致SCA数量减少(图7),这表明高浓度的IAA-Asp会对SCA生长和繁殖力产生不利影响。
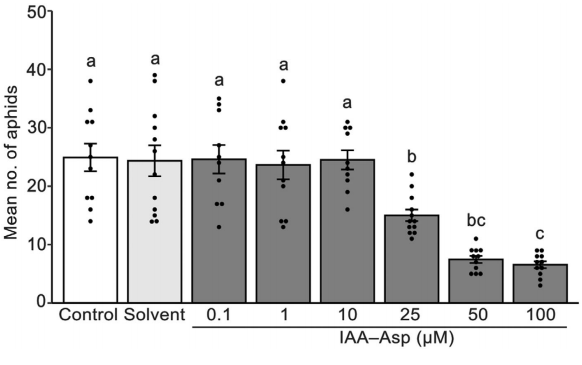
图7 不同浓度IAA-Asp处理对高粱蚜生长影响

作者利用高粱褐中脉(bmr)突变体来了解高粱对SCA的防御机制。与野生型相比,Bmr12功能缺失和Bmr12过表达(OE)分别增强了对SCA的抗性和易感性。对蚜虫取食行为的监测表明,与野生型和过表达材料相比,SCA在bmr12突变植株上到达首个筛管元素阶段(SEP)所需的总时间最长。转录组学和代谢组学分析表明,与野生型和过表达材料相比,bmr12突变植株的生长素代谢发生了改变,特别是IAA-Asp水平升高。此外,外源施用IAA - Asp恢复了Bmr12-OE植株的抗性,人工饲料蚜虫饲养试验生物测定显示IAA - Asp与对增强SCA的抗性有关。